Электроэнцефалограмма. анализ с точки зрения теории информации (начало)
07.02.2002
"ЭЭГ отражает функциональную активность мозга и, таким образом, зависит от степени организации мозговых систем".
Это цитата из книги читаемого и уважаемого автора многих учебников и серьезных научных трудов. Это положение - одно из основных и несомненных, изложено кратко и точно. В других книгах встречается модификация: "ЭЭГ отражает функциональное состояние мозга". С этим никто не спорит и сомнению не подвергает.
Если же попробовать уточнить эту аксиому, некоторые неясности начинаются со второго слова цитаты. Где отражает? И в чем? В зеркале, в мозгу исследователя, в компьютере? И как отражает, реализуя законы оптики, радиотехники или иные?
Если не останавливаться на этом неудачном термине и попробовать его заменить на "ЭЭГ описывает...?". Неприемлемо, т.к. количество информации в электроэнцефалограмме много меньше того, которое обрабатывает мозг за этот же период времени и разрешение метода и количество каналов записи не сопоставимо с количеством активных элементов мозга. Описывать функциональное состояние (или активность) мозга ЭЭГ, конечно же, не может.
"ЭЭГ коррелирует...?". Можно предположить, что это несколько ближе к точным терминам (и вычислению при необходимости может быть подвергнуто и стиль девятнадцатого века напоминает несколько менее). И нужно добавить: "иногда коррелирует", с учетом того, что существуют иные гипотезы.
Но, с точки зрения математика, занятого обработкой сигналов, неизбежен вопрос: "А с чем коррелирует электроэнцефалограмма? Необходимо определение понятия функционального состояния мозга".
Существует типичное мнение, что это само собой разумеется, и что такие вопросы могут задавать только дилетанты, которых к больному на пушечный выстрел подпускать нельзя. Возможно, эта гипотеза имеет свои обоснования, но до тех пор, пока не удается однозначно и математически корректно определить понятие функционального состояния головного мозга (или функциональной активности), этот термин придется, к сожалению, исключить т.к. не может быть использовано понятие, не поддающееся определению.
Поэтому необходимо дать определение самой электроэнцефалограмме, причем так, чтобы формулировка была однозначно понята математиками, специалистами по теории информации - всеми теми, кто реально занимается обработкой сигналов.
Предположим, что ЭЭГ - результат записи (здесь, несомненно, если «грамма», то запись) спонтанного электрического феномена живого головного мозга (достоверно известно, что после смерти мозга ЭЭГ исчезает), регистрируемого на кожных покровах поверхности головы (в отличие от электрокортикограммы) в диапазоне частот десятые доли - десятки герц и в амплитудных значениях единицы - сотни микровольт. Сигнал ЭЭГ имеет нулевое математическое ожидание (т.к. для регистрации используются усилители переменного тока) и близкий к нормальному закону распределения при исследовании реализаций, больших единиц секунд. Сигнал ЭЭГ нестационарен в большинстве реализаций и его осциллограмма напоминает смесь ограниченного по полосе розового шума с амплитудно-модулированными гармоническими составляющими с малой частотной девиацией.
Возможно, это определение не вполне соответствует врачебному, но имеет (наряду с недостатками) и свои преимущества т. к. термин "розовый шум", например, - четко определенное в технике понятие, однозначно описывающее спектр шумового сигнала, в отличие от термина "функциональное состояние мозга", который неизвестно что описывает.
Но как быть с главным вопросом? Какова связь ЭЭГ (или ее изменений) с деятельностью мозга? По этому поводу существует огромное количество гипотез, и диапазон оценок колеблется от "связи нет" до знаменитого высказывания Джибса "Электрическая запись, которую мы регистрируем, это язык самого мозга". Необходимо заметить, что и очень крупные специалисты позволяли себе выражать сомнение в связи ЭЭГ с функционированием мозга и высказываться как академик Анохин. (Может потому и позволяли, что крупные...).
Ничто, впрочем, не мешает сделать предположение, что правы были все исследователи. Но электроэнцефалограмма описывает не деятельность мозга, а результат работы только одной его системы. Системы удержания постсинаптического потенциала.
И в этом предположении есть, к сожалению, существенное отличие от общепринятой концепции, что электроэнцефалограмма коррелирует с деятельностью всего мозга, как структуры. А высказанная гипотеза предполагает, что система удержания постсинаптического потенциала имеет свои характеристики, свои принципы функционирования, вероятно, очень слабо связанные с системой обработки информации мозгом. Поэтому совершенно правы те исследователи, которые никакой связи между психической деятельностью (например) и ЭЭГ не находили.
В пользу того, что ЭЭГ не язык мозга, а результат работы некоторой системы, связанной не со всеми мозговыми функциями, можно привести простой аргумент: сигнал электроэнцефалограммы вышел на поверхность, т.е. распространился по мозговым структурам. Именно поэтому он не может быть суммой ВСЕХ тех сигналов, которые обрабатывает мозг, иначе пространственное разрешение внутри мозговых образований должно стремиться к нулю, а суммарный сигнал такого большого количества нейронов должен быть белым шумом (здесь следует оговориться, что пространственное распределение ЭЭГ не карта поля, а карта тока, что не всегда учитывают).
Естественно, что всякое предположение требует доказательств. Особенно с учетом того факта, что происхождение самой ЭЭГ вопрос до сих пор не вполне ясный.
Используем результаты известного эксперимента: изолированный участок коры головного мозга генерирует псевдогармонический сигнал с частотой около 2 герц с аддитивным шумовым компонентом. Спектральные характеристики этого сигнала отличаются от спектра ЭЭГ только смещением всех компонентов в область низких частот. Вопрос: что 'отражает' этот сигнал – функциональную активность изолированной коры? Какая может быть 'функциональная активность', если на входе и выходе устройства ничего нет, т.е. нет ни входов, ни выходов (они ведь отрезаны). Какие сигналы может обрабатывать этот участок, отключенный от всего. Думает о своей грустной судьбе изолированная кора? Здесь, конечно, приходят на помощь концепции самовозбуждения системы слабосвязанных осцилляторов, Уолтеровская 'стабилизация через флуктуации', разнообразные теории образования паттернов ЭЭГ (сканирование внешних данных) и т.п. Но, не пытаясь разобраться в гипотезах происхождения ЭЭГ, сделаем предположение: в достаточно большом изолированном участке коры автоматически включилась система удержания постсинаптического потенциала. И ничего она не обрабатывает, никакой функциональной активности не отражает. Этот сигнал свидетельствует только о том, что некоторая система работает. И у нее есть свои характеристики, свои законы функционирования и распространять эти закономерности на работу всего мозга не стоит, ведь изолированный от всего участок коры и функционирующий в полном объеме мозг отличаются принципиально. А спонтанные сигналы генерируют очень сходные.
Но, если предполагать, что ЭЭГ есть результат работы системы удержания постсинаптического потенциала, экспериментальные данные и гипотезы начинают весьма удачно укладываться в единую концепцию. В том числе и 'стабилизация через флуктуации', предложенная Уолтером (физики более удачно называют это стабилизацией системы методом принудительных осцилляций) - следует предполагать, что постсинаптический потенциал – опорное напряжение второго входа компаратора и динамический порог необходим для регенерации кодоимпульсных последовательностей, прошедших через реальный канал связи, т.е. получивших паразитную амплитудную модуляцию. Доказательство опубликовано неоднократно - любая запись любой последовательности внутриклеточных спайков демонстрирует разные амплитуды у разных спайков, т.е. кодоимпульсная последовательность подверглась паразитной (или не паразитной, как в принципе работы клеток Рэншоу) амплитудной модуляции. Так, впрочем, и должно быть, т.к. затухание в каналах связи существует, и синапс поврежденный может внести мультипликативную помеху, да мало ли может быть причин.
Теория Мак-Каллока тоже очень хорошо согласуется с этим предположением. Если частота альфа-ритма точно совпадает с максимальной частотой предъявления визуально распознаваемых объектов, т.е. альфа-ритм - кадровая развертка зрения - то эта частота и должна определяться частотой сигнала, поданного на один из входов устройства сравнения (иначе в системе возникнут биения и некоторые поданные объекты будут не только не распознаны, но и не все будут обнаружены). Но необходимо некоторое уточнение: частота предъявления объектов должна быть ВДВОЕ НИЖЕ частоты альфа-ритма, если частота альфа-ритма является частотой дискретизации (согласно основным положениям Найквиста). Если же альфа-ритм реализует временную апертуру дискретизации, то эквивалентная частота дискретизации соответствует фронту/спаду альфа волн, т.е. УДВОЕННОЙ частоте альфа-ритма - гипотеза приходит в соответствие с экспериментальными данными. При этом нужно учесть, что альфа-ритм относительно симметричен, и очень малая временная апертура квантования (как в технических устройствах) при малом быстродействии мозговых структур невозможна в принципе.
Если оценить гипотезу с позиций пейсмекерной теории ЭЭГ – увеличения медианной частоты спектров при понижении уровня пересечения мозга – можно считать, что пейсмекерная теория практически идеально описывает наращивание системы удержания постсинаптического потенциала и изменение ее основных характеристик.
Данные о том, что ЭЭГ продуцируется только двумя цитоархитектоническими слоями коры из шести - вполне уместны. Не могут ведь ВСЕ мозговые структуры заниматься только удержанием постсинаптического потенциала - это означало бы прекращение выполнения других функций.
Можно привести еще множество сопоставлений, но имеет смысл попытаться экспериментально подтвердить гипотезу 'ЭЭГ описывает работу системы удержания постсинаптического потенциала, а с остальными функциями мозга коррелирует настолько, насколько они связаны с этой системой '. Можно предложить следующий способ решения данной задачи: если бы в распоряжении экспериментатора был метод непрерывной оценки количества информации, содержащейся в электроэнцефалограмме и пациент с периодически возникающими существенными расстройствами мозговой деятельности и последующей ее нормализацией то:
-
Если во время пароксизмов будет зарегистрировано увеличение или уменьшение количества информации в ЭЭГ то - ни подтвердить, ни опровергнуть эту гипотезу невозможно.
-
Если удастся найти случаи, когда количество информации в ЭЭГ не изменяется во время пароксизмов, заведомо вызвавших существенное изменение деятельности мозга то - либо гипотеза верна, либо ЭЭГ вообще никак с деятельностью мозга не связана.
Понятно, последний тезис можно исключить, ЭЭГ с деятельностью мозга несомненно связана. То есть для подтверждения гипотезы необходимо найти такого больного и научиться оценивать количество информации в ЭЭГ (хотя бы относительное, об абсолютном не может быть и речи по причинам, обсуждение которых далеко выходит за пределы желаемого размера текста).
Больного такого найти несложно, а оценить количество информации труднее. Но, следует учитывать, что ЭЭГ многоканальное исследование. Пусть коэффициенты корреляции между любыми двумя каналами ЭЭГ равны нулю. Это означает, что все отведения ортогональны и сигнал ЭЭГ несет максимально возможное количество информации т.к. ЭЭГ в любом из отведений не может быть получена в результате любых линейных и нелинейных преобразований сигналов, зарегистрированных в любых других отведениях. Пусть коэффициенты корреляции между любыми двумя каналами ЭКГ равны еди нице. Это означает, что данные любого канала могут быть получены из лю бого другого и, фактически, многоканальная система не нужна, достаточно одноканальной записи, и сигнал ЭЭГ в данном случае несет минимально возможное количество информации, определяемое количеством информации в одном (или любом другом) канале. (Согласно положению теории информации - если форма сигнала известна заранее - сигнал информации не несет.) Реальные записи всегда представляют собой промежуточный случай между показанными двумя крайними вариантами, и оценить относительное количество информации можно просуммировав квадраты коэффициентов корреляции. В действительности, задача значительно сложнее: необходимо учитывать количество каналов регистрации, наличие аддитивного шума, всегда присутствующего в реальной записи и, самое главное, неортогональность системы отведений (что впрочем, для данной задачи не помеха - даже если система отведений существенно неортогональна - она все равно остается инвариантной во времени и смещение оценок будет одинаковым для всех экспериментов, т.е. данные между собой сравнивать можно).
Для оценки ортогональности многоканальной системы отведений (а в случае системы, инвариантной во времени, - относительного количества информации) применим метод, описываемый специально разработанной для этой цели формулой:
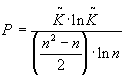 ,
,
где 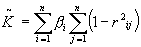
и с учетом наличия шумов реальной системы –
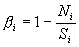 ,
,
где 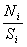 - соотношение шум/сигнал.
- соотношение шум/сигнал.
- соотношение шум/сигнал.
Разумеется n- количество каналов записи и r- коэффициенты корреляции.
Естественно, вычисления ведутся непрерывно со скользящим окном данных, длина которого определяется требованиями эксперимента.
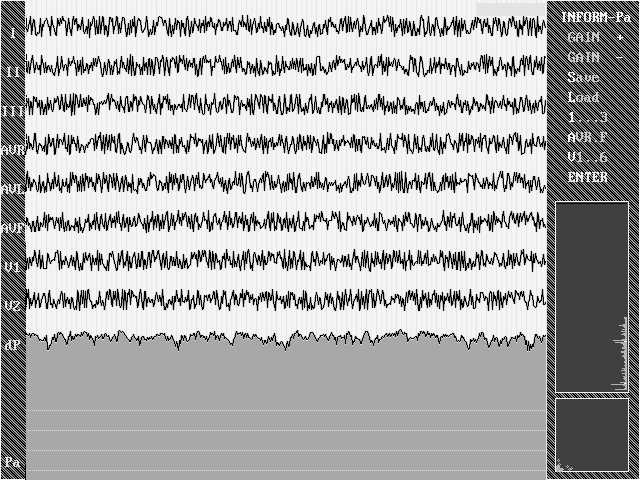
Рис.1.
Проверим программу: из формулы следует, что при подаче на вход системы белого шума параметр Р должен быть максимален и флуктуировать тем меньше, чем длиннее реализация (в данном случае шум представляет собой сигнал, совершенно некоррелированный в n-мерном пространстве, где n-количество каналов записи). Из иллюстрации 1 видно, что предположение подтверждается и Р все время находится вблизи верхнего предела (вопрос, почему он не достигает его, очень сложен и отвлечет от рассмотрения основных проблем. Достаточно будет сказать, что, при увеличении реализации, Р действительно асимптотически приближается к пределу, обусловленному качеством генератора случайных чисел в синтезаторе белого шума).
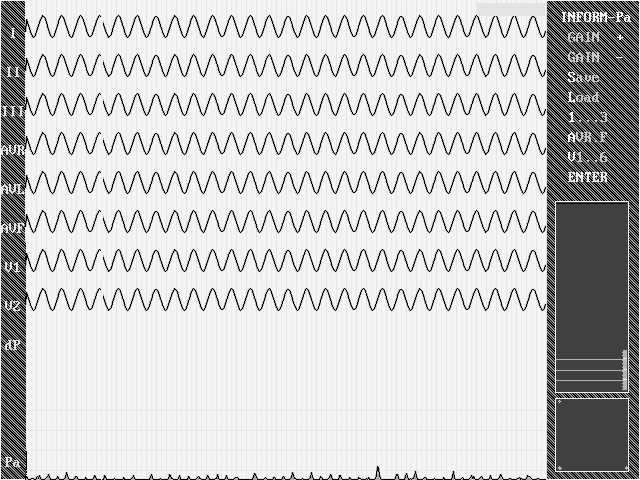
Рис.2.
Проверим программу также с помощью генератора синусоидальных колебаний. Очевидно, что при подаче на все входы системы любого гармонического сигнала, параметр Р должен устремиться к нулю вне зависимости от частоты и амплитуды колебания (ненулевые, хотя и близкие, значения параметра Р обусловлены в данном случае наличием шума квантования и погрешностями дискретной математики при вычислении коэффициентов корреляции: как видно из иллюстрации 2 эти погрешности крайне малы и проверяемая система готова к работе с реальными сигналами).
Проверим данные на более простом сигнале и попытаемся определить относительное количество информации в сигнале электрокардиограммы т.к. ЭКГ более 'понятный' т.е. сравнительно легко интерпретируемый сигнал, однозначно связанный во временной области с фазами сердечного цикла. Не будем забывать, кстати, что кардиохирурги по данным ЭКГ и электрокардиостимуляции делают (и успешно) свои операции, о чем электроэнцефалографистам и мечтать не приходится. И терминология у них однозначно определяет тип патологии (представим на минуту, что во время кардиохирургической операции физиолог начал пользоваться терминами типа 'паттерн, заинтересованность, ирритация, дезорганизация').
Итак - иллюстрация 3. Сигнал ЭКГ приводит к ОЖИДАЕМЫМ изменениям параметра Р. В этом случае длина скользящего окна данных равна 30 мсек. Видны сокращения предсердий, желудочков, изменение количества информации во время процесса реполяризации, хотя параметр Р во время диастолы должен вести себя иначе (он должен доходить до верхнего предела), так как во время диастолы сигнала ЭКГ нет, есть только шум усилителей и электродов. Причина техническая - в программе добавлен генератор очень низкоамплитудного гармонического сигнала (чуть выше уровня шумов усилителей). Это сделано для наглядности, чтобы случайные флуктуации параметра Р во время диастолы не мешали анализу его изменений во время систолы.
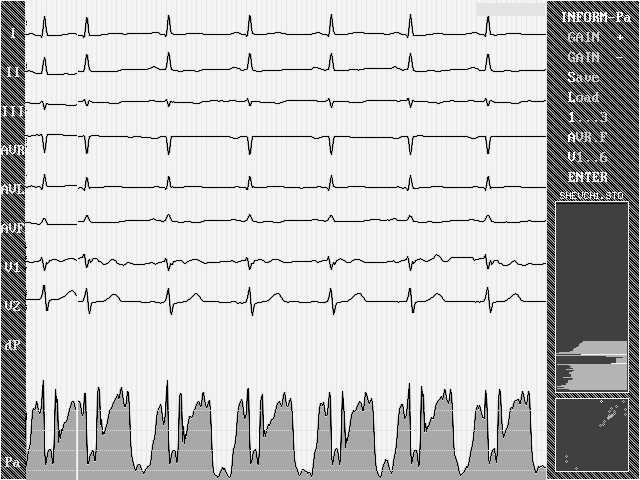
Рис.3.
Вернемся к анализу электроэнцефалограммы. Посмотрим на поведение параметра Р во время регистрации ЭЭГ здорового человека (Иллюстрация 4). При длине скользящего окна данных 30 мсек. (как и при любом другом коротком окне) наблюдаются колебания кривой, описывающей относительное количество информации, с частотой ВДВОЕ БОЛЕЕ ВЫСОКОЙ частоты основного ритма ЭЭГ.
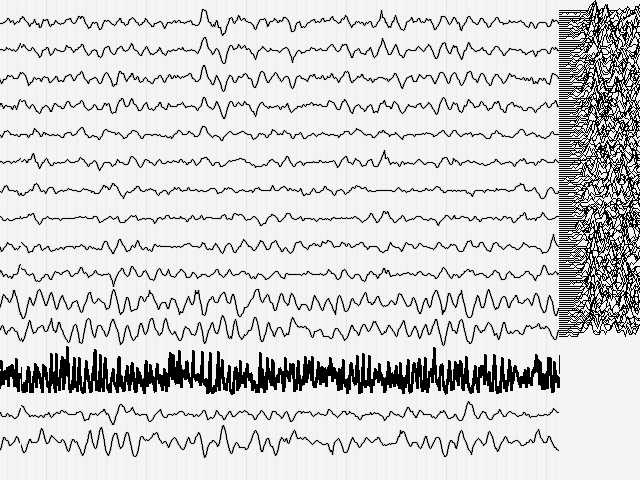
Рис.4.
Не будем делать из обнаружения этого феномена далеко идущих выводов, однако примем его к сведению. Естественно, что физиологическая интерпретация - удел специалистов, но очень похоже, что теория Мак-Каллока получает дополнительное экспериментальное подтверждение.
Увеличим длину скользящего окна (Иллюстрация 5). Высокочастотные колебания исчезли, однако теперь ничто не мешает оценивать относительно медленные изменения Р. Повторим задачу эксперимента: у больного с периодически наступающими существенными нарушениями функций мозга необходимо оценить поведение параметра Р.
Можно предполагать, что эпилептиформные разряды, соответствующие малому эпилептическому припадку у больного, не вызывающего диагностических сомнений при хорошей эффективности противопароксизмальной терапии и отсутствии сопутствующей патологии вполне удовлетворят требованиям эксперимента.
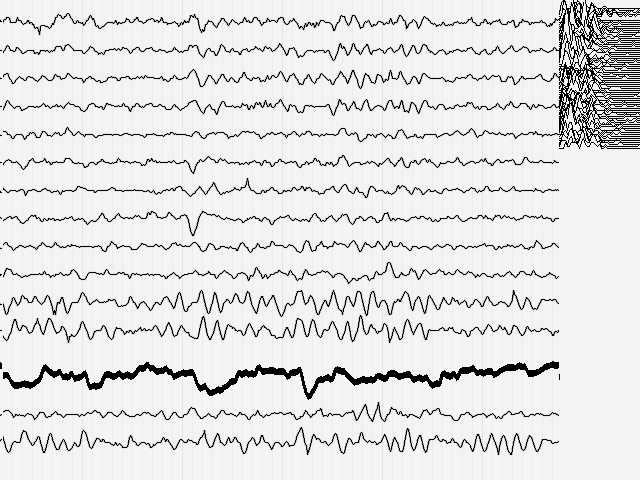
Рис.5
Действительно, во время малого припадка наступает существенное нарушение функций мозга, связанных с обработкой поступающей информации и управлением деятельностью других систем организма (нарушение мышечного тонуса и т.п.).
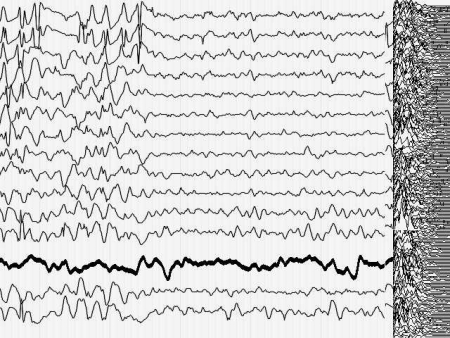
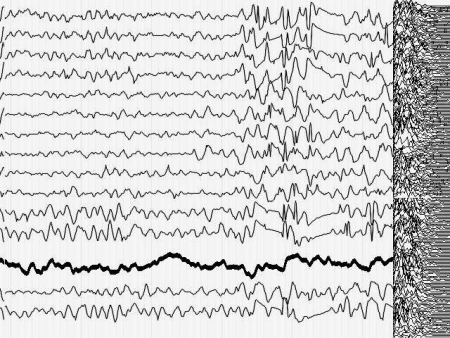
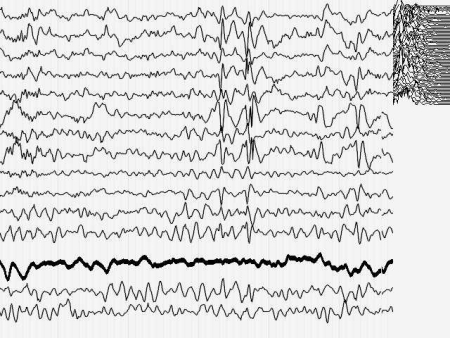
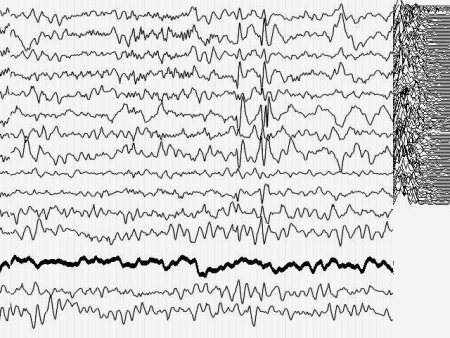
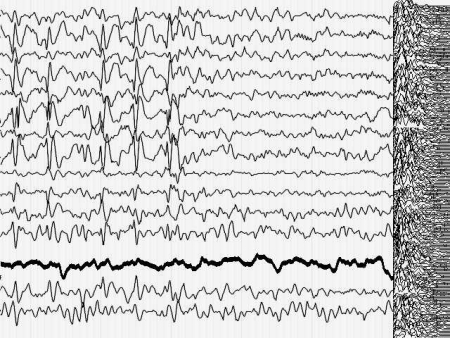
Кроме того, момент малого припадка сравнительно легко определяется по данным ЭЭГ. Для эксперимента необходимо оценить поведение кривой, описывающей поведение параметра Р до припадка, во время припадка и после него. Желательно также иметь повторяющиеся припадки, чтобы зарегистрировать систематические изменения Р и исключить элемент случайных совпадений. На иллюстрациях 6 представлены именно такие записи ЭЭГ.
Результат безнадежен: систематических, т.е. типичных изменений поведения параметра Р не выявлено как при визуальном просмотре, так и при оценке сжатых спектральных областей кривой, описывающей Р. Иными словами - количество информации в электроэнцефалограмме практически не изменилось (точнее - эти изменения неотличимы от спонтанных колебаний) во время разряда, который существенно нарушил функционирование мозга.
 |
 |
 |
 |
 |
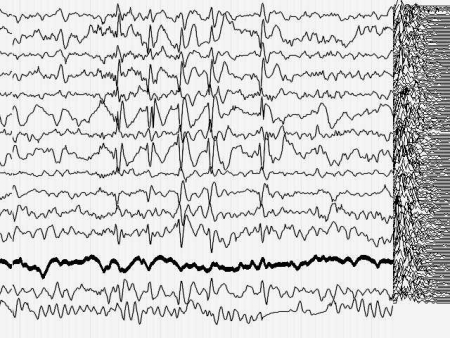 |
Рис. 6
Если бы ЭЭГ действительно существенно коррелировала с функционированием ВСЕХ мозговых систем - такой результат невозможен в принципе. Представим: электрическая запись работы некоторого исправного прибора несет такое же количество информации, как и запись работы прибора, временно вышедшего из строя. Конечно, этого быть не может, но ведь должно быть объяснение, почему припадок наступил, функции мозга нарушены, на ЭЭГ простым глазом прекрасно видны высоковольтные разряды, а информационный поток остался прежним.
Ответ может быть таким - ЭТО НЕ ТА ИНФОРМАЦИЯ, т.к. ЭЭГ описывает поведение только одной системы, которая подверглась воздействию. Да, в ней наблюдаются какие-то возмущения, она пытается самостабилизироваться, что мы и видим как высоковольтные разряды, но принципы ее функционирования остались прежними, катастрофы не произошло.
Сейчас можно задать вопрос: что это за информация 'та' и 'не та'? Почему используются такие ненаучные термины? Ответ прост - для наглядности. Дело в том, что оценка количества информации отнюдь не простое дело, очень мало напоминающее классическое определение количества бит при бросании монетки.
Если понимать информацию как меру устраненной неопределенности, то творчество графомана (или значительная часть содержимого сети Интернет, например), практически не несет информации, хотя объем этого творчества вполне может занимать очень много очень реальных мегабайт.
Вернемся, однако, к обсуждению результатов эксперимента. Еще раз уточним: при анализе электрокардиограммы сокращение предсердий однозначно вызывает систематически повторяющееся изменение количества информации, как и сокращение желудочков, и процесс их реполяризации (иллюстрация 3). А при анализе электроэнцефалограммы высоковольтный пароксизм НЕ ВЫЗВАЛ изменений, которые можно отличить от спонтанных флуктуаций.
А теперь вспомним Джибса: "Электрическая запись, которую мы регистрируем, это язык самого мозга". Можно, конечно, предположить, что существуют и такие языки, но если информационный поток ЭЭГ не изменился во время пароксизма, то с чем же мы имеем дело, что это за информация, поступающая по каналам электроэнцефалографа? И о чем свидетельствуют наши красивые кривые.
Посмотрим на запись ЭЭГ на иллюстрации 7. Попробуем сделать обычную визуальную оценку кривой. Можно сказать, что представлена ЭЭГ взрослого (потому что альфа-ритм сформирован и его частота составляет 9-10 герц) человека с закрытыми глазами (высокий уровень альфа-ритма, и он явно доминирует) в состоянии бодрствования (уровень медленноволновой активности мал, и альфа-ритм модулирован). Зарегистрирован также один высоковольтный гомолатерально-синхронный полифазный разряд в виде комбинации спайк-полиспайк волн с последующими низкочастотными относительно высоковольтными компонентами, очень напоминающими медленную волну.
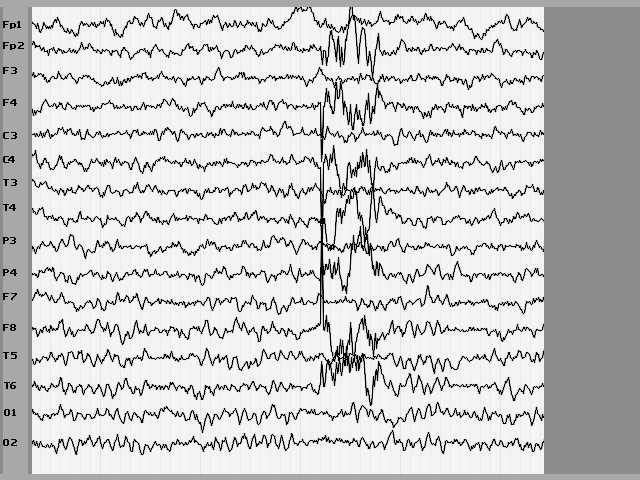
Рис. 7
Вероятно, в данном случае может идти речь об эпилептиформной активности. Что еще можно добавить? Не следует забывать, что истории болезни нет, о больном ничего не известно. Конечно, если бы иметь дополнительные данные в виде анамнеза, клиники, данных параклинических исследований ...
Но задача - визуально оценить ЭЭГ. Понятно, что если провести функциональные пробы и мониторинг ночного сна и фармакологические тесты, можно будет кое-что добавить.
Итак, добавлений и далеко идущих выводов делать не стоит, чтобы не попасть в неловкое положение, когда история болезни станет доступна.
И вот теперь стоит сообщить - ЭТО НЕ ЭЛЕКТРОЭНЦЕФАЛОГРАММА. Это, к сожалению, имитатор, используемый для некоторых специальных целей. Для доказательства приводится текст программы имитатора.
//ИМИТАТОР
{
if(j<21) //Для номера канала, меньшего 21
{a[j]=(a[j]*10+random(80)-40)/11; //Получаем розовый шум простейшим
// методом
a[j]+=sin(i1/50)*sin((double)i1/1.5)*j/4+random(5)-2;
//Добавляем амплитудно перемодулированные
//гармонические компоненты
//и аддитивный белый шум
}
if(j%2&&j<21) //Для нечетных каналов
{if(i1>320&&i1<380) a[j]+=random(31)-15;
//В определенный момент времени подаем
//высокоамплитудные случайные колебания
if(i1>320&&i1<322&&j%2) a[j]*=5;if(i1>322&&i1<325&&j%2) a[j]/=80;}
}
//КОНЕЦ ИМИТАТОРА
Здесь, пожалуй, стоит принести извинения за слишком вольный стиль изложения и невольную мистификацию и сообщить, что существуют значительно более совершенные имитаторы, работу которых даже очень опытный электроэнцефалографист от реальной записи отличить не может (проверено экспериментально).
Итак, нас вводят в заблуждение две синусоидальных функции и один генератор случайных чисел. Хорошо хоть о 'заинтересованности' диэнцефальных образований ничего не было сказано...
Хотя, вряд ли и в заблуждение. Сигнал в целом был оценен правильно: гармонические компоненты замечены, модуляция обнаружена, частота гармонического сигнала приблизительно определена, момент резкого повышения амплитуды не пропущен. А вот с шумоподобными сигналами дело явно хуже (возможно именно это - неустранимый недостаток визуальной оценки) и тот факт, что фаза гармонических сигналов во всех каналах неизменна (чего в принципе не может быть в электроэнцефалограмме) отмечен не был. И еще - в лучших традициях электроэнцефалографической семиотики 'альфа-ритм' был назван модулированным, хотя в действительности имеет место перемодуляция. Точно так называют модуляцией перемодуляцию в ЭЭГ - эта ошибка мешает правильному пониманию того, что генераторов альфа-ритма не один, а множество и их биения и дают картину перемодуляции. Впрочем, где теория единого генератора, там и до эквивалентного диполя недалеко.
Но, возвращаясь к основной теме, уточним: что в этом имитаторе дало нам полезные сведения о его устройстве (пусть имитатор станет на короткое время черным ящиком)? Синусоидальные компоненты - раз, их модуляции - два, импульсное повышение амплитуды - три и специфическая форма некоторых компонентов - четыре. С шумом дела плохи - неясно шум ли это, либо сложная смесь сигналов (что, впрочем, не удивительно - кто видел осциллограмму работающего факс-модема - знает, картина ограниченного по полосе шума и ничего более, а информация там перекачивается килободами).
Как же произошла оценка полезной информации? По-разному: анализировались видеоимпульсы (в случае высоковольтных пароксизмов) и радиоимпульсы (в случае оценки альфа-ритма и его модуляций). И такая оценка используется во всей клинической ЭЭГ - работа идет то с видео, то с радиоимпульсами (в отличие от оценки ЭКГ, где четко и жестко (но и субъективно) оценивают только видеоимпульсы, да еще и с дополнительной синхронизацией - импровизированной временной привязкой).
И в таком подходе нет ничего удивительного, т.к. механизмы генерации ЭЭГ до конца не понятны, а принципы анализа складывались исторически по мере обнаружения новых феноменов и сопоставления их с клиникой. А предложение радикально изменить подход к анализу электроэнцефалограммы неприемлемо вследствие огромного накопленного экспериментального материала и опыта ее клинической оценки. Поэтому всякие новые методы обработки должны быть совместимы с клинически опробованными. Но вопрос об информативности самого метода ЭЭГ остается пока неясным. И, вполне понятно, что эксперимент с оценкой количества информации во время пароксизмов не может помочь определению степени значимости каждой волны ЭЭГ или любых групп волн, т.е. можно сказать только о том, что ЭЭГ коррелирует с деятельностью НЕ ВСЕХ мозговых систем. А предположение о связи ее с системой удержания постсинаптического потенциала так и остается гипотезой.
Можно, разумеется, признать эксперимент бессмысленным, а сделанные выводы неверными. Но попробуем оценить результаты 'ad absurdum', т.к. единственное, в чем можно не сомневаться - корректность вычислений и исправность аппаратуры. Зададим вопрос: какой должна быть система, чтобы количество информации в канале связи (который мы перехватываем) не изменилось при ее временном частичном повреждении (или другом нарушении функционирования)?
Как ни странно, с точки зрения теории информации ответить на этот вопрос просто - система должна иметь нулевую избыточность в канале связи. При этом, разумеется, сигнал становится шумоподобным (что и наблюдается в случае электроэнцефалограммы). Простейший пример: у факс-модема периодически отключается входное устройство, а модулятор продолжает работать, используя в качестве входных данных шум. И при любом анализе сигналов телефонной линии мы не обнаружим моментов частичного выхода прибора из строя.
Пока не дешифруем сам сигнал. Тогда и станет ясно, что осмысленные данные периодически прерываются. Может быть, и в проведенном эксперименте наблюдался такой эффект? Может быть действительно, каждая волна ЭЭГ несет некоторое дешифруемое сообщение? И ЭЭГ - в самом деле язык мозга?
Маловероятно. Кроме шумоподобного характера сигнала (но аналогия не доказательство) ничто не говорит в пользу такого вывода. А возражений много.
Начиная с того, что электроэнцефалограмма, вероятно, не запись перехвата канала связи, т.к. это суммарные данные электрической активности большой группы источников сигналов, да еще и с низким пространственным разрешением и с вкладом шунтирующего эффекта внемозговых образований. Кроме того, спектр ЭЭГ не совпадает по виду со спектром ограниченного по полосе белого шума, т.е. избыточность канала связи нулевой быть не может. И флуктуации количества информации во время эксперимента наблюдались, да еще и с разной периодичностью, что противоречит гипотезе о полном использовании пропускной способности канала связи. А спектр имеет явный вклад компонентов вида 1/f, т.е. похожих на фликкер шумов, что, видимо, не следует игнорировать.
Вообще, вопрос о вкладе собственных шумов нейронных структур не рассматривается (насколько мне известно) в гипотезах генерации электроэнцефалограммы мозгом. Но, всякая система, в которой есть перемещение электрического заряда, шумит по физическим причинам и не шуметь не может. И если вкладом тепловых, дискретных (дробовых) шумов на частотах регистрации ЭЭГ можно пренебречь, то исключить вклад фликкер шума - нельзя. Действительно, имеется система, включающая в себя около 1.5*10^9 элементов с тонкими мембранами, в которой циркулируют сигналы амплитудой сотни микровольт - единицы милливольт. Более того, известно, что нейроны при отсутствии входных сигналов имеют довольно значительный потенциал поляризации.
Такая система, даже при отсутствии передачи электрических импульсов от нейрона к нейрону, ДОЛЖНА генерировать фликкер шум, уровень которого будет прямо пропорционален потенциалу поляризации и обратно пропорционален средней площади изолирующего перехода, т.е. мембраны нейрона, причем коэффициент шума будет зависеть от величины флуктуаций носителей заряда на поверхностях мембран.
Следует учитывать, что даже незначительный уровень шума, поступающего на один из входов компаратора в аналоговом устройстве (нейроне) может и должен приводить к высокому уровню мультипликативной помехи в выходном сигнале, особенно при работе в амплитудных областях, близких к порогу. Иными словами - выходная последовательность импульсов будет промодулирована шумом и при демодуляции выходного сигнала (интегрировании в простейшем случае) шум будет восстановлен, причем уровень его станет выше исходного и будет определяться коэффициентом усиления системы (и характеристиками демодулятора).
Если же учесть, что в нейронных структурах действительно обнаружены цепи реверберации с малым коэффициентом затухания, влиянием шума тем более нельзя пренебрегать. Действительно, достаточно одному нейрону кратковременно отреагировать на случайный шумовой сигнал и вся система обеспечит его циркуляцию до некоторого критического момента стробирования (или запоминания, согласно некоторым из теорий). Понятно, такая система вряд ли могла развиться в процессе филогенеза, да и вообще, скорее всего, неработоспособна. Но без шумовых компонентов неясно, какие сигналы транслируют нейронные сети в условиях отсутствия внешних импульсов: получается абсурдное устройство - ревербератор тишины. Тот факт, что некоторые нейроны генерируют спонтанную активность, не отменяет необходимости учета шума, т.к. генераторы с внутренним возбуждением, в роли которых выступают эти нейроны, также трансформируют свой внутренний шум в фазовую нестабильность выходного сигнала (в более общем случае это положение сводимо к тому, что невозможно получить абсолютную спектральную чистоту гетеродина).
Разумеется, так упрощенно подходить к гипотезам происхождения ЭЭГ нельзя, но и гипотезы генерации электроэнцефалограммы должны учитывать некоторые физические ограничения – например, неизбежности вклада шумовых компонентов в работу любой аналоговой системы.
Но все же интересно, каким образом в процессе эволюции была решена проблема шума, ведь перехода к полностью цифровым системам не произошло и структура нейрона мало отличается у очень разных организмов, т.е. решение было найдено на довольно ранних стадиях развития многоклеточных.
Остается один путь решения проблемы шума - сужение полосы пропускания сист ем методом повышения добротности частотно-избирательных цепей. Естествен ным следствием такого решения становится необходимость фазовой демодуляции как обеспечивающей наилучшие характеристики систем обнаружения и распознавания сигналов (экспериментальные работы в области электрофизиологии действительно обнаруживают такой механизм).
Понятно, что с учетом неустранимого вклада шумовых компонентов, системы нейронных сетей должны быть достаточно точно настроены на определенные частоты и высокая добротность этих сетей определяет псевдогармонический характер генерируемых ими колебаний. В случае отсутствия входных сигналов и выполнения ЛИВ условия (т.е. отсутствия механизма параметрической генерации), выходной сигнал нейронной сети должен определяться характером шума на входе и ее амплитудно-частотной характеристикой.
За этим может последовать вывод о том, что если ЭЭГ результат аддитивной суммации выходных сигналов множества нейронных сетей, она должна определяться псевдогармоническими компонентами наиболее высокодобротных сетей и псевдошумовыми менее добротных, т.е. можно синтезировать сигнал со спектром, идентичным спектру ЭЭГ данного пациента, если в качестве исходных данных использовать шумовой сигнал его поляризованных клеток. При этом потребуется лишь адаптировать амплитудно-частотные характеристики трактов передачи сигналов (т.е. имитаторов нейронных сетей). Естественно, абсолютно безразлично, как будут спроектированы эти имитаторы. Существенным является лишь характер шума, обусловленный флуктуацией носителей заряда на поверхности мембран.
Безусловно, можно получить, например, сигнал, соответствующий по спектру и внешнему виду электроэнцефалограмме данного пациента из шумовых компонентов его электрокардиограммы во время диастолы. Однако не следует забывать, что аналогия - не доказательство и подобных цирковых трюков достаточно в электрофизиологических работах. Анализ моделей нейронных сетей также никого ни в чем не убеждает, т.к. применяя нелинейные преобразования можно получить что угодно из чего угодно. Тем не менее, попробуем получить сигнал, сходный с электроэнцефалограммой из шума, имеющего существенный вклад мультипликата, используя только комбинации высоко и низкодобротных квазинейронных сетей (т.е. оставаясь в рамках предложенной гипотезы).
На иллюстрации 8 видно, как из мультипликата белого шума белым шумом (график 1) получается весьма напоминающий ЭЭГ сигнал (график 2), состоящий из суммы сигналов 3 и 4, причем сигнал 4 - результат работы высокодобротной системы с постоянной времени 0.1 сек и шумовой девиацией 0.03 сек, а сигнал 3 - результат суммирования нескольких систем с малой добротностью и разными постоянными времени.
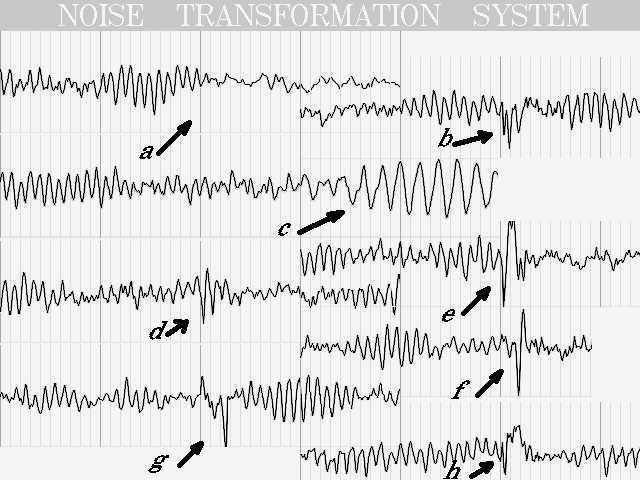
Рис. 8
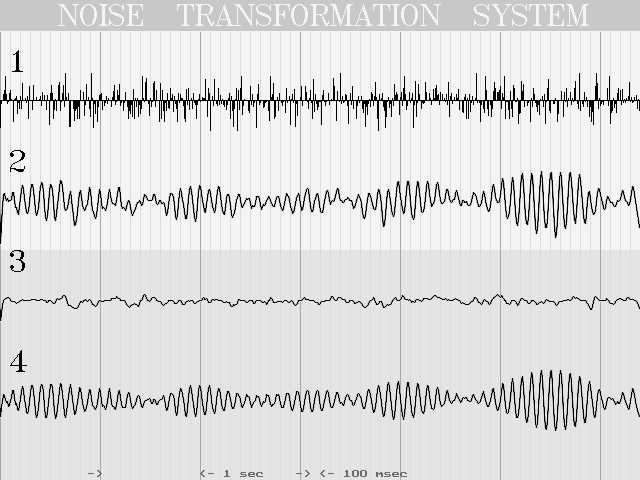
Рис. 9
Еще более интересные результаты получаются, если вводится произвольное управление одним параметром системы. Из иллюстрации 9 следует, что можно получить картину депрессии альфа-ритма(a), полиспайки, (b) медленноволновую тета-активность (c), заостренную альфа-волну (d), пик-волну (e), двуфазную острую волну (f), однофазную острую волну (g) и пик-медленную волну (h).
Причем все эти феномены получаются, если менять длину кольца, по которому происходит распространение сигнала, или обеспечивать дополнительное замыкание кольца в произвольной точке (что полностью соответствует механизму re-entry). А входным сигналом во всех случаях был мультипликат шума. Понятно, что после получения красивой картинки, возникает соблазн объяснять все патологические феномены ЭЭГ механизмом re-entry. Но, вероятнее всего, он имеет существенно вторичное значение, т.к. тех же результатов можно добиться параметрическим управлением нейронной сетью. Действительно, достаточно предположить, что в зоне опухоли, например, время отклика нейрона на входное воздействие возрастает (вследствие парабиоза) и эквивалентная частота настройки системы понижается при сохранении добротности - результат: генерация медленноволновой активности. Кроме того, нейронные сети не могут быть неуправляемы, а единственный способ управления такими системами - параметрический. Впрочем, при объяснении происхождения спайков и острых волн без механизма re-entry, может быть обойтись и не удастся.
Интересным и важным результатом эксперимента является тот факт, что характер шума на входе имеет тем меньшее значение для формы выходного сигнала, чем больше высокодобротных цепей включено в систему (в полном соответствии с постулатами теории информации, кстати). Иными словами - не суть важно, какой шум присутствует на входе - белый, розовый или голубой. И не имеет значения, что подается - мультипликат или чистый шум. Более того, несущественно шум ли это или псевдошумовой сигнал. Главное, чтобы на входе не было гармонических сигналов или их смеси с целочисленно-кратными частотами.
В общем, итоги хорошо согласуются с данными экспериментальных электрофизиологических работ и полностью противоречат современным постсоветским гипотезам происхождения ЭЭГ с их "громадными нейронными ансамблями", "спайковой бомбардировкой", функциональной активностью мозга и прочими чудесами.
Но, преследуя вполне прагматическую цель - обосновать корректные принципы анализа ЭЭГ - необходимо все-таки сформулировать какую-то хоть и упрощенную гипотезу формирования сигнала электроэнцефалограммы.
Итак, с позиций теории информации (хочется надеяться, что правильно понимаемых), механизм возникновения сигнала ЭЭГ, вероятнее всего, обусловлен:
-
Существованием кольцевых нейронных сетей, настроенных на определенные частоты и перестраиваемых параметрически механизмом управления, находящимся вне пределов нейронного кольца.
-
Характер генерируемых этими сетями выходных сигналов практически полностью определяется параметрами цепей и очень мало зависит от обрабатываемых мозгом сигналов.
-
Хотя эти сети и предназначены для обработки информации, их выходной суммарный сигнал не может быть дешифрован, т.к. производимое ими преобразование сигналов не является обратимым (т.е. с помощью ЭЭГ невозможно узнать, какие сигналы и как обрабатывались).
-
Каждая из этих сетей должна быть непрерывно связана с механизмом управления, находящемся в глубоко расположенных мозговых образованиях (вероятнее всего, кольцо сети замыкается в них, иначе невозможно объяснить, почему расположенная ниже корковых образований опухоль способствует генерации медленноволновой активности).
-
Характер сигналов, циркулирующих по этим сетям - псевдошумовой, причем в случае спонтанной ритмической активности шум трансформирован в джиттер сигналов или в частотную девиацию.
-
Каждое проявление изменений корковой активности, регистрируемое на ЭЭГ должно иметь корреляты в глубоко расположенных мозговых образованиях, т.е. не может быть только коркового очага эпилептиформной активности, например. Последнее положение, к сожалению, принципиально расходится с общепринятой точкой зрения.
Разумеется, все эти выводы следует отнести к системе регистрации ЭЭГ, не использующей принципы пространственной фильтрации и не оценивающей фазовых взаимоотношений между сигналами в разных каналах записи.
Если вернуться к оптимизации принципов анализа электроэнцефалограммы, то следует признать, что эмпирически разработанные методы ее оценки далеко не плохи. Следует лишь четко разделить случаи, когда необходим анализ радиоимпульсов, т.е. ритмов и когда видеоимпульсов, т.е. единичных специфических феноменов. И в любом случае не имеет смысла оценивать группы волн как последовательность осмысленных сообщений и объединять распознаваемые единичные феномены в группы и подвергать их анализу, предназначенному для оценки радиоимпульсов.
Но наиболее существенным выводом, применительно к анализу ЭЭГ, будет положение о том, что анализ псевдошумовых низкоамплитудных компонентов электроэнцефалограммы бесперспективен, т.к. это результат суммации сигналов множества нейронных сетей (в том числе и высокодобротных), и извлечь информацию не удастся, т.к. спектры сигналов перекрываются и сигналы существуют одновременно. Что впрочем, практикующие физиологи давно используют без всяких теоретических обоснований.
А принцип паттерна, видимо, при таком подходе сформулировать не получится.
Возможно, при других способах оценки, при использовании методов анализа, предназначенных для существенно нестационарных сигналов или эффективных методов распознавания образов, его и удастся (или уже удалось) формализовать.
Возможно, впрочем, имеет смысл, оставив нездоровый оптимизм моделей нейронных сетей, вернуться к по-прежнему непонятному сигналу электроэнцефалограммы. Нездоровый потому, что существует множество моделей генерации ЭЭГ, более или менее остроумных. И моделей нейронов и моделей сетей достаточно и все они вполне могут генерировать сходный с ЭЭГ сигнал. Но аналогия - не доказательство и точный ответ о конструкции мозга будет сформулирован тогда, когда будет получена его принципиальная схема в статике и динамике (не более, не менее). А пока можно спорить, верна или нет модель Вилсона-Коуэна, строить предположения о том, как может быть устроен мозг, и обсуждать гипотезы.
Но не стоит использовать принципы теории информации, при оценке вариантов типа "А может быть это так?". Значительно более продуктивным подходом представляется оценка типа "Как не может быть" и "Как не может не быть". Например, дискретные элементы системы НЕ МОГУТ НЕ ШУМЕТЬ, а вся система НЕ МОЖЕТ БЫТЬ спроектирована так, чтобы этот шум не учитывать и не компенсировать (разумеется, если она работоспособна). И не может быть, чтобы в процессе филогенеза и копии его в онтогенезе (т.е. по мере наращивания системы) было потеряно управление филогенетически более молодыми областями, если существуют промежуточные варианты работоспособных систем (либо необходимо отменить теорию эволюции). А это значит, что не может быть изолированно-коркового очага эпилептической активности, как ни противоречит это маститым теориям. И даже экспериментальный корковый очаг можно "выключить", если использовать только воздействие на глубоко расположенные структуры. Вопрос этот до конца не выяснен, но так не может не быть. И такие механизмы можно назвать и противоэпилептической системой и рассмотреть понятие эпилептического порога и т.д. и все это будет корректно до тех пор, пока гипотеза будет предусматривать управление снизу вверх (параметрическое, вероятнее всего).
Впрочем, это большая задача, требующая отдельного рассмотрения. Пока имеет смысл ограничиться единичными феноменами электроэнцефалограммы, т.е. рассматривать их как видеоимпульсы, причем каждый отдельно, игнорируя для упрощения марковские компоненты процесса.
Для любого анализа видеоимпульсов их необходимо сначала обнаружить и распознать. Задача обнаружения и распознавания - общее место при анализе сигналов (в радиолокации, радиоперехвате и т.д.), но в области анализа ЭЭГ часто можно встретить случаи распознавания без обнаружения и еще более странные процедуры. Проблема заключается в том, что единичные феномены обнаруживаются и распознаются чаще всего визуально при неясных решающих правилах. И одному доктору кажется, что эта острая волна выделяется из альфа-ритма, другому не кажется, третий считает, что это артефакт, и мнений в итоге столько и они так полярны, что можно разлагать их в ряды.
Зададим вопрос - где начинается и где заканчивается пик-волна? Нет, это не само собой разумеется, это очень даже не простой вопрос, ведь если начало (вследствие высокочастотных компонентов пика) легко обнаруживается, то окончание процесса уходит в спонтанные псевдошумовые компоненты ЭЭГ. Если необходимо экспериментальное подтверждение этого тезиса, можно сделать несколько ксерокопий пик-волны (одной и той же) и попросить электрофизиологов проанализировать. В общем случае, разброс оценок моментов начала и конца пароксизма будет тем больше, чем выше степень маститости экспертов. Причем один будет оценивать начало от момента пересечения изолинии, другой от предыдущей волны, а третий по каким-то своим критериям, которые понять не удастся.
А как правильно? Как, в соответствие с успехами в области обнаружения и распознавания цели (и поражения, иногда), провести правильное обнаружение патологического феномена (принимая во внимание, что пик-волна - феномен не менее важный, чем неопознанная цель над нашей территорией).
Можно попробовать сформулировать ответ с позиций "как не может быть". Не может быть определения начала феномена с момента пересечения изолинии, т.к. любой аддитивный низкочастотный компонент или тренд могут вызвать существенную ошибку во временной области. Не может быть определения по изменению спектральных компонентов сигнала, т.к. при уменьшении реализации нарастает неопределенность в частотной области (принцип неопределенности пока никому отменить не удалось).
И не помогут неклассические методы спектрального оценивания, адаптивная фильтрация, ничто не поможет.
Допустимым решением при априорном знании особенностей ЭЭГ сигнала будет определение начала и конца интересующего патологического феномена по предыдущему и последующему экстремумам функции, описывающей сигнал (априорные предположения базируются на том факте, что между экстремумами ЭЭГ- сигнала мало что меняется, и межэкстремальную область хорошо описывает нелинейная интерполяция, что подтверждается существованием эффективных ЭЭГ-компандеров, работающих именно таким способом).
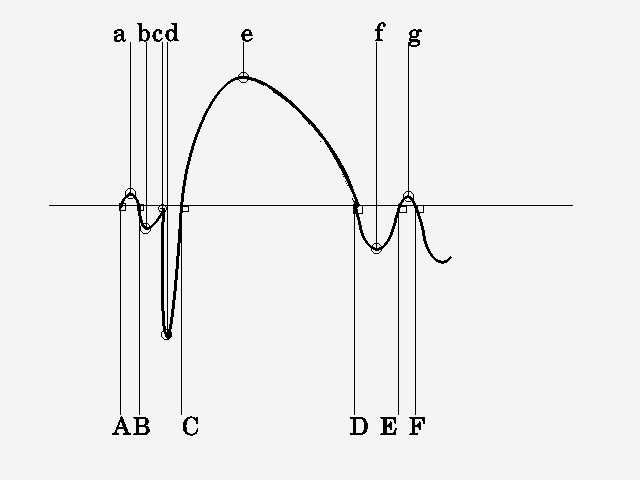
Если решение допустимо (хотя, возможно, и неоптимально) - логично будет остановиться именно на нем т.к. вариантов можно предложить великое множество и доказательство чем один лучше другого - тема отдельного исследования. Так на иллюстрации 2.1 показаны варианты обнаружения пик-волны по пересечениям изолинии (временной интервал B-D) и по экстремумам (временной интервал c-f). Очевидно, что обнаружение по пересечениям нулевой линии дает "неправильный" результат, а вариант с экстремумами соответствует типичным врачебным представлениям о том, где начинается и где оканчивается пик-волна.
Распространим принцип обнаружения видеоимпульсов по экстремумам для всех феноменов ЭЭГ (в этом случае необходимо будет объявлять феноменом каждую волну - от пика до пика). И, хотя метод не вызывает принципиальных возражений, его необходимо модифицировать для решения задачи автоматического обнаружения феноменов. Действительно, если использовать этот принцип в заявленном виде
 |
 |
|
Рис.2.1
|
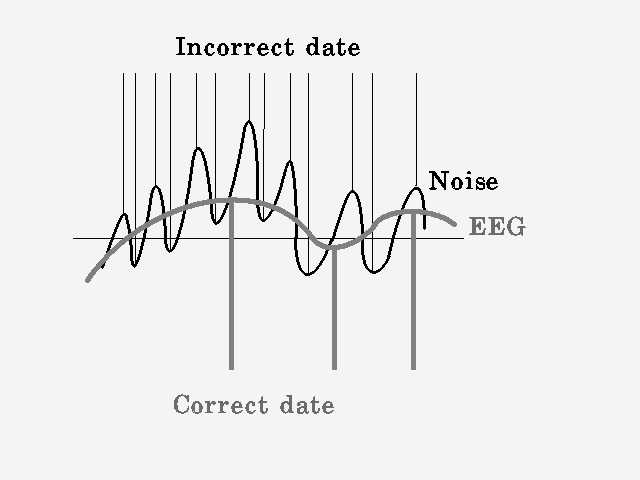
Рис.2.2
|
(рис. 2.2), ничего, кроме самых высокочастотных компонентов проанализировано не будет (а феномены в группы объединять нельзя). Поэтому придется обеспечить "завал" амплитудно-частотной характеристики примерно на 6 db/октаву и фильтрацию сетевой помехи (чем больше, тем лучше). Такое решение не является оптимальным, но вполне допустимо и дает в случае анализа ЭЭГ удовлетворительные результаты (с подобной проблемой столкнулись разработчики систем уплотнения линии в телефонии и успешно ее решили с помощью анализа группы волн, но в нашем случае группы недопустимы).
Будем считать, что задача обнаружения экстремумов в сигнале ЭЭГ успешно решена (и это довольно близко к истине), сетевая помеха полностью подавлена, и исходная электроэнцефалограмма может быть точно восстановлена глобально заданным кубическим сплайном по координатам экстремумов (следует оговориться, что здесь нет ничего нового, подобная задача неоднократно успешно решалась при обработке и компрессии сигнала ЭЭГ).
Будем считать также, что обнаружены ВСЕ феномены ЭЭГ, причем начало и окончание каждого - точно известны. Можно, понятное дело, оставить это утверждение на совести автора, но не будем забывать, что оно не хуже любого другого. Подходит время решать задачу распознавания.
Впрочем, существующее положение дел в электроэнцефалографии таково, что вряд ли кто-либо рискнет утверждать о полном решении задачи корректного распознавания всех феноменов ЭЭГ.
Допустим, однако, что последовательность видеоимпульсов может быть дешифрована (в простейшем случае - классифицирована). В качестве примера используем телевизионный сигнал: любое его изучение, спектральный, корреляционный анализ, трансформация в фазовую плоскость и т.п. не могут помочь понять содержание телевизионной передачи. Да, будут найдены закономерности кадровой и строчной развертки, обнаружена высокая корреляция между строками, получен спектр и еще много хороших, правильных и интересных (обнадеживающих, как сейчас говорят) наблюдений. Но что передают - можно будет понять лишь тогда, когда будет обеспечена развертка сигнала на экране. Не будем забывать также, что развертка в плоскости экрана сопровождается свертыванием во времени в пределах кадра (для зрительного восприятия).
Попробуем и в случае экстремумов ЭЭГ получить изображение процесса (здесь лишь совпадение терминов - никакого соответствия изображению при некоторых специальных математических преобразованиях нет) (рис. 2.3). Очевидно, что для любого ЭЭГ сигнала можно получить некоторое изображение в системе координат: логарифм дифференциальных времен / логарифм абсолютных значений дифференциальных амплитуд экстремумов. Понятно, что ЭЭГ сигнал после такого преобразования восстановлен быть не может. Очевидно также, что изображение немодулированного гармонического сигнала будет представлять собой точку, угловая модуляция вызовет трансформацию изображения в горизонтальную линию, амплитудная - в вытянутый по вертикали эллипс. Т.е. любые изменения в амплитудных значениях будут приводить к увеличению размеров фигуры по вертикали, а изменения частоты - по горизонтали.
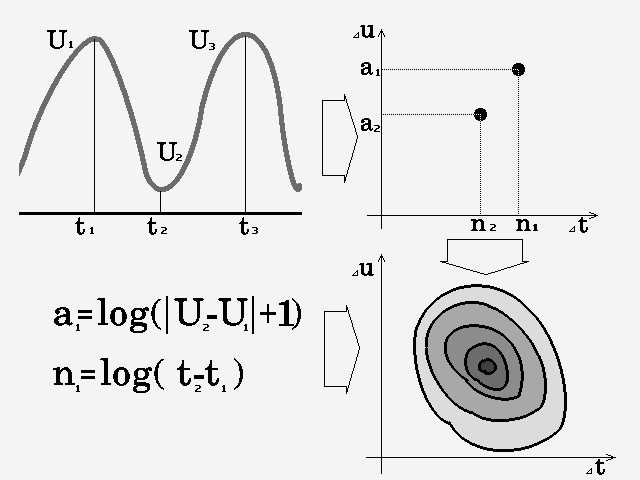 |
 |
|
Рис.2.3
|
Рис.2.4
|
Таким образом, каждая волна ЭЭГ имеет свои координаты на плоскости, обусловленные ее амплитудой и длительностью. Если волны ЭЭГ идентичны по этим параметрам и отличаются только формой в межэкстремальной области - они займут одну и ту же точку. Следствие - после этого преобразования синусоидальный сигнал от пилообразного или меандра отличить невозможно.
Однако полезным свойством такого преобразования после анализа достаточно большой реализации будет возможность не только оценить процесс по амплитудным и частотным параметрам, но и сопоставить характеристики каждой волны с характеристиками процесса. Например: есть единичный спайк, который отличается малой длительностью и высокой амплитудой - разумеется, его координаты будут существенно отличаться от координат процесса и его легко РАСПОЗНАТЬ, что и требовалось. Если есть группы спайков,- они образуют свое изображение, отстоящее на плоскости от основного. Конечно, это утверждение относится не только к группе спайков, но и к любой волне или группе волн, характеристики которых хотя бы по одному параметру отличаются от основного процесса (здесь явно появляется призрак паттерна).
Категории:
- Войдите, чтобы оставлять комментарии
|
|
|
Комментарии
Костя
18.05.2016 - 14:10
Постоянная ссылка (Permalink)
Здравствуйте, что означает диагноз ЭЭГ: выраженный невротический паттерн ээг усиления процессов синхронизации коркового электрогенеза?